DONATE
MEMBERSHIP
Menu
Animals
Plants
Wildlife Care
Saving Species
Wild and Fun
News
Oliver Ryder Ph. D.
Safeguarding Biodiversity and Preserving Hope
October 16, 2023
Surprised by Science
November 18, 2021
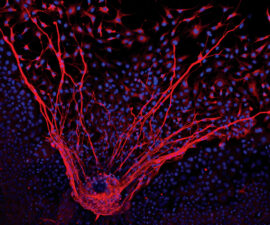
Stem Cells to the Rescue
November 23, 2020
Recommended
The Hatch of 2020
Wildlife Care